


The Nature of Nature
Turkheimer E. (2015). The nature of nature. In Philosophical Issues in Psychiatry III: The Nature and Sources of Historical Change, ed. KS Kendler, J Parnas, pp. 227–44. Oxford University Press.

Purchase HERE
When we speak of genetic transmission, we are addressing the real problem of how far differences in the genotype and differences in the environment contribute to the development of such a character…. If genetic differences make a contribution it is important to try to find out what we can about their nature. Gottesman and Shields, 1962, p.10.
Nineteent Century: The Galtonian Paradigm
Questions of nature and nurture, famously, originated with Charles Darwin’s cousin, Sir Francis Galton, who borrowed the phrase from Prospero’s description of Caliban:
A devil, a born devil, on whose nature Nurture can never stick; on whom my pains, Humanely taken, all, all lost, quite lost. And as with age his body uglier grows, So his mind cankers. I will plague them all, Even to roaring."
Galton (1875), like so many who came after him, liked the sound of the phrase “nature-nurture:”
The phrase 'nature and nurture' is a convenient jingle of words, for it separates under two distinct heads the innumerable elements of which personality is composed. Nature is all that a man brings with himself into the world; nurture is every influence without that affects him after his birth. (p. 9)
There can be no doubt that questions of nature and nurture scratch a deep itch in the human psyche. Although most discussions of nature and nurture eventually turn into deconstructions, it is worth considering seriously for a moment how strong our nature-nurture intuitions are. Children are not given lessons to learn how to run or to speak their native language: they just learn it, and even our nagging awareness that children raised swaddled and isolated might not learn to do either does not dissuade us from thinking of language and locomotion as “inborn,” “innate,” or (in the modern era), “hard-wired.” But no one, or almost no one, emerges full-blown as a virtuoso oboe player or champion pole-vaulter: those we do have to take lessons for. We say they are learned, the result of specialized and systematic exposures. Dogs bark, but they don’t roll over unless you teach them how.
Galton’s intuition about nature and nurture went one step beyond the classification of abilities into the inborn and learned, however. It is not only running in general that feels like it is inborn, it is also the fact that some people can run faster than others. Humans differ on even the most universal of behavioral characteristics, and it seems-- still referring only to widely-held intuitions, perhaps misguided but worth taking seriously for a while-- that some of those differences are more the result of nature and others the result of nurture. Few of us would contend that we might have been concert musicians or Olympic champions if only we had the training[1], but getting to Carnegie Hall is at least to some extent a matter of practice. Spelling out the intuition in this way has already clarified one important conclusion: nothing is purely the result of either nature or nurture. Olympic sprinters have to train, great oboe players have some hard-to-define quality called musical talent, and even the ordinary among us can improve ourselves with practice.
Reading Galton 150 years later is an experience in simultaneously appreciating the breadth of his genius and the opacity of the blinders placed on him by the common prejudices of his era. In his famous studies of "eminence," he examined the familial clustering of men (again, accepting without question that there was no place else to look) of great accomplishment in Victorian society. Along the way he invented much of what we recognize as modern social science, starting with a compulsion to quantify what he studied, and including the correlation coefficient and, as we will see in a moment, at least a portion of the twin method.
It comes as no surprise today that Galton’s men of eminence were, indeed, clustered in families; what is remarkable is the extent to which Galton was willing to accept this clustering as evidence of "nature." Although Galton labored mightily to develop methods to differentiate nature and nurture, from a modern perspective it is clear that Galton was so firmly rooted in the pre-genetic, casually racist and rigidly stratified socioeconomic confines of the nineteenth century that he could not fully appreciate the magnitude of the fundamental problem. It is more obvious today that clustering of human beings in families is the result of both genetic relatedness (which Galton recognized as "nature") and environmental relatedness of various kinds, including especially social class, the elephant in the room throughout Galton's studies.
Galton was, however, not so thoroughly naive. As pointed out in Rende, Plomin and Vandenberg’s (1990) excellent history of the early development of the twin method by Galton and others, in a 1876 article entitled “The History of Twins, As A Criterion Of The Relative Powers of Nature And Nurture,” Galton considered the role of the environment in familial clustering and differentiation. Galton had some premonition of the existence of identical and fraternal twins, but did not have the scientific tools to fill it out. As a result he did not use the twin method in what we now think of in the “classical” way, via comparison of the similarity of monozygotic and dizygotic twins. Instead he analyzed the developmental tendency of twin pairs to become more alike or different over time, an idea with strikingly modern resonances of its own (Beam & Turkheimer, 2013).
Twentieth Century: Twin Method
The development of the classical twin method as we know it today awaited genetic advances by Weinberg at the turn of the twentieth century, leading to an explosion of twin studies in the twenties, mostly on the subject of human intelligence (Rende, Plomin & Vandenberg, 1990). The classical twin method is based on the existence of two distinct types of twins: monzygotic (MZ), who share 100% of their DNA sequence (within an order of approximation; epigenetics and other aspects of contemporary genomics have complicated this simple picture somewhat), and dizygotic (DZ) who are essentially ordinary siblings, sharing on average 50% of the DNA sequence, but who happen to share a womb and be born at the same time.
The conceptual, genetic and statistical details of the twin method have been described and debated many times already, and we will do so as briefly as possible here, with a goal of neither supporting nor undermining its many genetic and scientific assumptions. There are two sources of resemblance in twin models: a broadly genetic source, labeled A for Additive (about which more in a moment) and one environmental source, labeled C for common. We will have more to say about both of these, but for now it is simply an observation that members of twin pairs resemble each other because they share DNA on the one hand (A) and share a rearing environment on the other (C). The genetic source of resemblance is twice as large for MZ pairs than it is for DZ pairs, the family environmental source is the same for both of them. The third term, labeled E (for environment, though it is actually one of two environmental terms in the model), is not a source of resemblance: in fact it represents all the environmental factors that make siblings and twins raised in the same family different from each other. Given a sample of identical and fraternal twins that varies in some outcome, one can use these relationships to partition the variation in outcome into independent source of variation attributable to A, C and E. Before reviewing how such twin studies have come out for commonly studied forms of psychopathology, it is necessary to describe some details that are crucial to understanding the results.
The name additive that is given to the genetic term in the twin model is very important. What is being added in such models are the tiny effects of many individual genetic loci. A major task of genetics during the first half of the twentieth century was the unification of Mendelian genetics, in which characteristics like smooth and wrinkled peas “segregated” across generations in known ratios according to laws of discrete genetic transmission, with quantitative genetics, in which continuously distributed characteristics like height are transmitted from parents to children but do not segregate into types. Twin designs are very much an example of the latter. The integration is accomplished by assuming a very large number of segregating genes of small effect, which, when summed (thus the A) produce a quasi-continuous bell-shaped curve according the the binomial theorem.
An additional complication of quantitative models in psychiatric genetics is that for better or for worse, psychiatric outcomes have historically been represented as classifications rather than as continua. This adds one more layer to models of familial resemblance: variation in many discrete genetic loci is combined with whatever environmental risk can be identified to create a latent continuous distribution of liability to disorder with a threshold: individuals with liability scores greater than the threshold develop the disorder. The quasi-categorical character of psychiatric disorder also means that twin similarities are usually expressed as concordances (percentage of pairs with one affected individual in which the other is also affected) rather that the correlation coefficients developed by Galton for the analysis of continuous traits like height and ability.
Also in the category of fortunately or unfortunately is the fact that twin and family studies result in an index, called a heritability coefficient, that varies from zero to one, expressing the proportion of variance in outcome that is accounted for by variance in genotype. The heritability coefficient is one of the most reified, vilified, and generally misunderstood parameters in the history of science, and its strengths and weaknesses do not need to be rehashed here. The current author has addressed the issues quite recently (Turkheimer & Harden); more (Visscher, Hill & Wray, 2008) and less (Charney, 2012) sanguine views are also available.
In any event, in the 1920s the twin method evolved from the early sketches provided by Galton to the “classical” method in which the similarities of identical and fraternal twins were compared and modeled. Although the slow and shifting development of reliable diagnostic systems and the relatively smaller sample sizes imposed by the rarity of psychiatric disorder may have resulted in somewhat more variable results than were obtained for more easily measured normal characteristics like intelligence, from an historical birds-eye view the results were unanimous. Psychopathology is heritable, in all its forms, in the simple sense that MZ twin concordances are consistently higher than DZ twin concordances; there is little evidence for systematic environmental effects originating in families; but there is persistent evidence of unsystematic environmental variability within families, which is simply a way of saying that identical twins raised together are substantially less than perfectly similar for any form of psychopathology. Everything is partially heritable and nothing is completely so: schizophrenia in its various taxonomic forms; bipolar and bipolar depression; later, the same was found to be true for anxiety and personality disorder, as those disorders were clarified with the development of official scientific systems of classification like the DSM.
The discovery that all major forms of psychopathology were substantially heritable was, at the time, a source of some degree of triumphalism among psychiatric geneticists (Seymour Kety, 1974, replying to Thomas Szasz’ (1960) Myth of Mental Illness, famously quipped, “If schizophrenia is a myth, it is a myth with strong genetic component.”) The ubiquity of genetic variance and the defeat of traditional family environmentalism led to a widely-held point of view that major psychopathology had turned out to be “genetic,” and that it would therefore only be a matter of time before the scientific details were filled in, as they had been for Huntington’s disease after the identification of…. Schwarz, for example, in the 1991 Review of Psychiatry, put it as follows:
Even as the epidemiologic [i.e. twin and family] studies continued, it was clear to the field that if there was a genetic etiology to schizophrenia, it would act through a biological mechanism. Therefore, studies aimed at understanding the biological pathophysiology of schizophrenia take on a new importance for genetic hypotheses, especially if the factor being studied is likely to be under somewhat simple genetic control (e.g. an enzyme level of neurotransmitter receptor density). (p. 85)
It was common at the time to go as far as to declare that schizophrenia is a “brain disease” (Henn & Nasrallah, 1982; Johnson, 1989; Heinrichs, 1993).
Chua and McKenna (1995), for example, stated,
In recent years, the hypothesis that schizophrenia is a biological brain disease has spectacularly gained the upper hand. The origins of this paradigm shift can be traced directly to two incontrovertible findings: first, the hereditary contribution to the disorder and, secondly, the antipsychotic effect of neuroleptic drugs. (p. 563)
To be fair, Chua and McKenna expressed at least a modicum of skepticism about the idea. What they shared with every other theorist who introduced the idea, however, is that they didn’t stop to analyze exactly what it meant for something to be a brain disease, or the role that genetic variance might properly play in the determination.
At the time, findings of genetic variance in psychopathology also sparked considerable resistance and controversy. From a contemporary perspective it may be difficult to remember that fifty years ago, the goal of behavior genetic studies of psychiatric illness was not so much to uncover the biological etiology of psychopathology, but simply to “unravel” (as was commonly said) genetic from environmental sources of etiology. The prevailing criticisms of the day were psychoanalytic and family environmental, holding that family dynamics were the most important determinant of psychopathology, including for schizophrenia (Lidz & Blatt, 1983). Although twin-study based behavior genetics still has its critics, are there any surviving “environmentalists” as regards psychopathology? In the middle of the twentieth century, the behavioral sciences were dominated by two great schools of thought that were, in different ways, deeply environmental in outlook: psychoanalysis and behaviorism. There are, of course, still psychoanalysts and behaviorists, but modern Freudians and Beckians don’t bother to deny the importance of genetic factors in mental illness; it is considered a given, an etiological background against which any treatment modality does its work.
The environmentalists-- the Laingians, the Szaszians, the humanists, the Sullivanians-- may been routed, but there were nevertheless worrisome signs that the Galtonian victory of nature over nurture in psychopathology still faced difficult obstacles. The first was the very ubiquity of heritability. It wasn’t as though the landscape of psychopathology had been subdivided into meaningful sections, some strongly genetic and others more environmental. Everything was heritable, and to more or less the same intermediate degree. To this day there is no well-documented theory of psychopathology showing that some forms are more heritable than others. (Much of this difficulty is based on foundational difficulties in the concept of heritability itself. See Turkheimer & Harden, in press, for a review.)
Even more potentially troubling, the ubiquity of heritability turned out to extend beyond the bounds of major psychopathology. Following from Galton, differences in human intelligence were found to be heritable; so were differences in normal personality. Moreover, differences in human traits that seem well beyond the usual extent of biological explanation are heritable, indeed no less heritable than schizophrenia or anything else. Political attitudes are heritable (Hatemi, Funk, Medland, Maes, Silberg, Martin & Eaves, 2009), religiosity is heritable (Waller, Kojetin, Bouchard, Lykken & Tellegen, 1990), how much television people watch is heritable (Plomin, Corley, DeFries & Fulker, 1990). And, in an example this author has pursued theoretically and empirically for the last twenty years, divorce is heritable (d’Onofrio, Turkheimer, Emery, Harden, Slutske, Heath, Madden & Martin, 2007). The heritability of divorce has implications for too-easy inferential leaps from quantitative genetics to genetic and biological explanation.
A third difficulty in the transition from quantitative genetic to mechanistic accounts of psychopathology is that as it became more and more possible to identify actual genes that caused psychopathology, it became increasingly clear that it was going to be very difficult to do so. It is commonplace to locate the beginnings of the search for DNA related to psychopathology with the completion of the Human Genome Project, but more limited searches for linkage with genetic markers has been possible for much longer than that, and the troublesome patterns that have come to characterize much modern genomic research-- early reports of large effects greeted with great enthusiasm, followed by disappointment in replication and meta-analysis-- plagued the field from the beginning. In a passage entitled, “The Pond is Empty,” Crow (2008), recalled
Many in linkage research thought that success was inevitable – one would ‘drain the pond dry’ and there would be the genes ! However, the reality is that in spite of a plethora of well-hyped findings no linkage claim has proven robust. In each case an apparent finding in a modest-sized population of families that was then used to ‘identify’ a candidate gene has not been found linked in more systematic and larger studies. (p. 1682)
In fact, the absence of genes of large effect for psychopathology should not have come as a surprise. As I have already described, the additive action of many genes of small effect is an assumption of routine quantitative genetics. When comparisons of polygenic and of single gene or “oligogenic” (specifying a handful of genes of intermediate effect against a polygenic background) models were actually conducted, they invariably favored the polygenic theories. Thoughtful psychopathologists of the era were troubled by the prospect: what kind of biological mechanism could be invoked if genetic effects were so completely distributed across genes? Faraone and Tsuang (1985) put it as follows:
The mode of inheritance has substantial implications for etiological research and clinical practice. A conclusive demonstration that a single major locus is involved in schizophrenia would hold the promise that a relatively direct biochemical pathway accounts for the psychophysiology of the disorder. If a multifactorial polygenic model describes the mode of transmission, the search for a simple biochemical pathway is likely to be less fruitful, (p. 44)
A final indication that the victory of nature of nurture was not complete is that the opposition never conceded. In fact, the growing biogenetic consensus of the time was paralleled, then as now, by an anti-hereditarian opposition that dismissed the entire quantitative genetic enterprise as ill-conceived. At the time, the most comprehensive attack on the foundations of behavior genetics is a book entitled "Not in Our Genes," by Lewontin, Rose and Kamin (1984). In the first pages of the book, the authors were clear about the twin foundations of behavior genetics they wished to undermine. The first is reductionism:
The first is reductionism-- the name given to a set of general methods and modes of explanation both of the world of physical objects and human societies. Broadly, reductionists try to explain the properties of complex wholes-- molecules, say, or societies-- in terms of the units of which those molecules or societies are composed. (pg. 5, emphasis in original)
The second is determinism:
The second stance is related to the first; indeed, it is in some senses a special case of reductionism. It is that of biological determinism. Biological determinists ask, in essence, why are individuals as they are? Why do they do what they do? And they answer that human lives and actions are inevitable consequences of the biochemical properties of the cells that make up the individual; and these characteristics are in turn uniquely determined by the constituents of the genes possessed by each individual. (pg 6, emphasis in original).
Lewontin, Rose and Kamin were correct in identifying reductionism and determinism-- as opposed to heritability-- as the important theoretical issues underlying nature and nurture. In their concluding chapter, they endorsed a view of biological systems according to which complexity of biological determination is the source of what we perceive as our psychological freedom from biology:
What characterizes human development and actions is that they are the consequence of an immense array of interacting and intersecting causes. Our actions are not at random or independent with respect to the totality of those causes as an intersecting system, for we are material beings in a causal world. But to the extent that they are free, our actions are independent of any one or even a small subset of those multiple paths of causation: that is the precise meaning of freedom in a causal world. When, on the contrary, our actions are predominantly constrained by a single cause.......we are no longer free. (pg. 289).
Although the authors of "Not in our Genes" recognized that heritability has an oblique relationship to their legitimate concerns about reductionism and determinism, they never gave up the attack on its empirical validity. Heritability, however, is precisely a means for computing genetic effects summed over a large "subset of those multiple paths of causation." In the kind of complex hierarchical biological model they endorse, heritability is not a problem to be explained away; rather, it is inevitability, an assumption of the model.
Lewontin et al.'s analysis of the genetics of schizophrenia is particularly telling: Although they have laid the groundwork for a compelling argument that the non-zero heritability of schizophrenia does not commit us to a biological "cause" of schizophrenia, any more than the heritability of divorce must lead to a neuroscience of marital status, they never make the case. Instead, they fall back on the traditional anti-hereditarian strategy of sniping at the methodology of twin and adoption studies, apparently hoping to convince the reader that a methodologically correct study (which, of course, is never actually conducted) would show the heritability of schizophrenia to be negligible. Finally, having convinced themselves that their stance against determinism commits them to radical environmentalism, they end the chapter with an endorsement of the views of Michel Foucault and R. D. Laing.
We can see in this progression the same confusions that plague traditional behavior genetics and biological psychiatry, precisely the confusions that Lewontin et al. set out to avoid: confounding of determinism and heritability, and commitment to a dichotomous view of the role of biology in behavior. Setting out to bring down reductionism, they end up in a futile attack on the empirical status of heritability; and once they have rejected reductionist models of schizophrenia, all they have their disposal is the opposite extreme, the discredited Laingian contention that schizophrenia is no more than a lifestyle, and a heroic one at that.
21st Century: The Nature Debate
In Turkheimer (2001) I wrote that “The nature-nurture debate is over.” The debating hasn’t actually stopped, of course, and in that sense my declaration was premature, but that isn’t really what I meant. Although there was obviously no particular date when it occurred, at some point around the turn of this century, the Galtonian nature-nurture project reached its logical conclusion: there is genetic variance in everything, psychopathology included; there is also environmental variance in everything; and most important, genetic variation does not imply genetic mechanism. The nature-nurture community in general, and the psychopathology community in particular (because questions of mechanism are especially important for psychopathology) have been trying to recover from this last conclusion ever since. The nature-nurture debate has not so much ended as it has been transformed into a debate about the meaning of the “nature” term in Galton’s “convenient jingle.” Call it the nature debate.

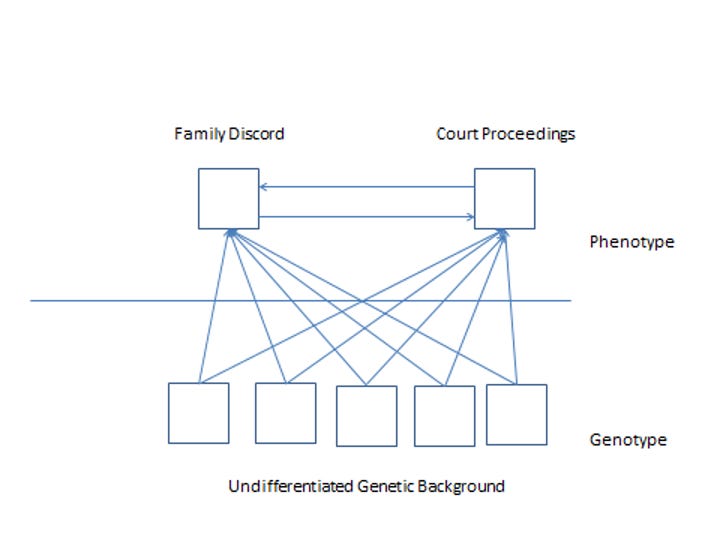
Figures 1 and 2
Establishing what the nature debate is about necessitates beginning with the obvious. Not all genetically-influenced characteristics are etiologically alike. Figures 1 and 2 illustrates the relationship between genetic structure and behavior for two “syndromes” that are observable at the level of behavior. Figure 1 represents Huntington’s Disease. A clinical neuropsychologist might observe that a subset of patients in a neurological clinic displayed a characteristic pattern of disparate symptoms. Among other symptoms, these patients display rapid cognitive decline in midlife, and a particular form of motor disturbance. Indeed, these clinical characteristics are what were observed by Huntington, well before the genetic basis of the disorder was known. Huntington’s Disease is “genetic” in two senses. The first is the Galtonian sense, according to which differences in the expression Huntington’s status are correlated with genotype and therefore cluster in families, but that is the less important part of what we mean when we say Huntington’s is genetic. Once the genetic nature of the disorder was known, our understanding of its etiology was changed. In particular, explanations at the level of behavior-- why people with particular kinds of motor disorders might be motivated to experience cognitive decline, or vice-versa-- became pointless. Both are caused by a single dominant gene. Differences in Huntington’s status contain genetic variance, and Huntington’s disease is explained by a genetic mechanism: these are two different things.
Figure 2 displays a similar set of circumstances for divorce. A social worker might observe a “syndrome” in which two kinds of activities co-occur in some individuals: they argue a lot with their partners, and spend time in court hearings. These two behaviors co-occur because the individuals in question are divorced, and the heritability of marital status ensures that genes are correlated with the likelihood that a person will have these experiences. Like Huntington’s, differences in marital status are correlated with differences in genotype. But in the case of divorce, we have not discovered a gene responsible for the relationship, or a well-specified mechanism linking the genetic and behavioral levels of explanation; presumably we don’t expect to discover one. Nevertheless, the heritability of divorce ensures that as married people descend into conflict with their spouses and head into court, their genes will come along for the ride. Nature, in the Galtonian, genetic epidemiological sense, contributes to both Huntington’s disease and divorce, but the nature of Huntington’s disease is different than the nature of divorce. The difference between Huntington’s disease and divorce is not that the former involves genetics and the latter does not; it is that the former is defined by genetics whereas the latter is defined by behavior.
A conventional account of the difference between Huntington’s disease and divorce would emphasize that Huntington’s is the result of a single major locus whereas divorce is polygenic, and that is of course true. One way to understand the problem faced by modern complex genomics is to say that we are still searching for coherent ways to specify genetic etiologies of the radically polygenic causal architectures that have turned out to be the norm for behavior. This problem has faced the field of psychiatric genetics from its earliest days. Gottesman and Shields’ (1962), in their classic theoretical and empirical account of the genetics of schizophrenia, were the first to insist on a polygenic model of inheritance, but they remained troubled by the relation between variation and etiology. In particular, they struggled with the Meehlian concept of “specific genetic etiology.” I have argued for some time that “specific genetic etiology” is exactly what we are talking about: Huntington’s disease has a specific genetic etiology, and divorce does not. Gottesman and Shields were convinced that their polygenic model of the transmission of schizophrenia was compatible with a specific genetic etiology, but Meehl (1962) himself was not. In his lengthy (and generally admiring) afterword to the volume, he wrote,
“One understands fairly clearly what it means to conjecture that a “big-effect monogene” is the specific etiology of a disease...But once we have excluded that simple situation, the very meaning of the phrase “specific etiology” begins to “fuzz up,” and I wish the authors had said more about how they conceive it.”(p. 376)
It is an extraordinary fact that fifty years later, biological and genetic psychiatry still lacks a coherent theory of mechanism that allows us to distinguish clearly between Huntington’s disease and divorce. There are, in my view, only three broad possibilities for how a theory of specific etiology of polygenic characteristics might work. The first two are the perpetually embattled doppelgangers that have dominated the discussion for a long time: reductionist hereditarianism on the one side, and radical anti-hereditarianism on the other.
The hereditarian view, which is not often spoken out loud but is, I suspect, held in the breach by many working behavioral and psychiatric geneticists, is that despite our intuitions to the contrary, it is only a matter of time before things like divorce succumb to mechanistic genetic analysis. The unstated assumption of biological reviews is that anything detectably syndromal, and certainly anything both syndromal and heritable, must have a genetic mechanism to explain it. This assumption is often adopted out of either explanatory desperation (If there is no genetic mechanism, how can science proceed?) or a misguided and oversimplified materialism (see, for example, Guze’s famous question, “Can psychiatry be too biological?” and my affirmative reply (Turkheimer, 1998) reply.) The second possibility is a long anti-hereditarian tradition holding that the Galtonian, quantitative genetic empirical work showing marital status to be heritable is methodologically flawed and biologically oversimplified, and that the failure to identify genetic mechanisms of complex behavior is simply a reflection of those mistakes. These arguments have survived, virtually unchanged, into the genomic era (e.g., Joseph, 2004).
The perpetual opposition between the reductionists and the anti-reductionists is potentiated by a common assumption that there are only two choices: something is either a brain disease or a lifestyle. If mechanistic genetic explanation cannot be brought to bear on psychopathology, the reductionists envision their field undone by naïve dualism, or at best, relegated to murky domains of social science. The anti-reductionists envision a dehumanized world in which all human suffering, up to and including marital status, has been reduced to a deterministic world of genetic effects. But fortunately for all concerned, it is actually glaringly obvious that most forms of psychopathology are neither brain diseases nor lifestyles. What is lacking is a sensible theory that offers some middle ground. The remainder of this paper will begin to outline such a theory.
The third option, that of developing a meaningful theory of biological and psychological explanation in psychiatry, will require giving up on the first two possibilities in a way that the poles of the old nature-nurture argument have never been able to do. Has there ever been a heritable syndrome for which the biogeneticists have finally said, “You know what? This syndrome may have some genetic variance like everything else, but it doesn’t have a genetic mechanism. We are going to have to leave this one to the psychologists.” Conversely, has there ever been a syndrome for which the anti-hereditarian community has, finally, conceded that despite omnipresent environmental variation, psychological explanation isn’t leading anywhere? The first task of any meaningful theory of biogenetic and psychological explanation is to distinguish them; no theory is plausibly independent of its mindless or brainless prejudices (Lipowski, 1989) unless it can offer a meaningful example of a syndrome that does not fit the author’s preferred explanatory modality .
My own account of the relationship between genetic vs. environmental variation on the one hand and biological vs. psychological explanation on the other is in its early stages. In Turkheimer, Horn and Pettersson (in press) we suggest that the hallmark of genetic variation that does not play a specific mechanistic role is that the multivariate structure of the genetic variability is indistinguishable from the structure of the phenotype. We build the case that human personality is the first well-documented example of a well-defined domain of behavior that is demonstrably heritable in the Galtonian sense but has nevertheless resisted all forms of genetic explanation for so long that the time has arrived to declare it “phenotypic.”
For example, our fundamental understanding of phenotypic differences in human personality have been developed with factor analysis. Large matrices of personality responses are analyzed with the goal of identifying the minimum number of latent dimensions that account for their multivariate structure. These exercises have led to the Five Factor Model of personality, which has become the almost universally accepted standard in the field. If one has access to genetically informative personality data on a similar matrix of personality items, it is possible to conduct the factor analysis separately on the genetic, shared environmental and nonshared environmental matrices. This possibility leads to an interesting question: if the phenotypic variance in personality can be described in terms of the Five Factor Model, what about the genetic variance? Perhaps if we factor analyze the genetic variation separately from the environmental, we can learn something about the genetic etiology of personality that contributes to the observable phenotype. But despite many attempts, the answer is that when we factor analyze the genetic variance-covariance matrix, we get-- the Five Factor Model. There is usually not enough shared environmental data to conduct a meaningful factor analysis, but if we factor analyze the nonshared environmental variance, we get-- the Five Factor Model again. John Loehlin, at the end of a thirty-year program of research on the topic, concluded, “… the primary organization of personality is at the level of the phenotype...” (Loehlin & Martin, 2013; p. 766).
Space does not permit a full adumbration of all of the ways in which the genetics of human personality looks like phenotypic human personality, described at a different level of analysis. Several things are worth noting briefly. First, attempts to specify the molecular genetics of personality, initially with linkage and candidate gene studies, and more recently with genome wide association studies (GWAS), have been singularly unsuccessful. As one would expect based on a radically polygenic, phenotypically driven model, there are associations to be found between personality and individual genes, and if sample sizes become large enough we will “detect” them at whatever level of statistical significance we like, but that is irrelevant to the question at hand. A rejection of the phenotypic null model requires discovery of genetic-level structures that display meaningful differences from phenotypic structure, in the way that (in the extreme case) the highly structured familial segregation of Huntington’s differentiates it from other forms of dementia or apraxia.
By the same token, the failure to develop a biologically interesting molecular genetics of personality should not be taken of evidence that the original impression of genetic variation in personality was incorrect. The two findings are compatible; in fact they are both specific predictions of a radically polygenic phenotype-driven model. Finally, characterization of the phenotypic hypothesis as “null” is important. Null hypotheses can’t be proven, in the sense that it is not possible to prove that a mechanistically meaningful personality gene will never be found. This is one of the reasons that dreams of biological explanation die so hard. All I can do is predict that in the long run personality will be seen to be phenotypic, make the case that things certainly look that way so far, and remind those who continue to expect a biological explanation to emerge that the burden of proof is ultimately on them.
The most comprehensive account of a post nature-nurture model of psychiatric explanation is found in Kendler (2013). Kendler begins with the universal heritability of “human traits and disorders,” which he accepts as evidence that, “because genetic effects must be expressed through biological processes, these findings suggest that psychiatric disorders are ‘biological.’” Later, however, Kendler is appropriately skeptical about this “weak” claim of biologicial explanation, because, as he correctly recognizes, in its most general sense the epithet “biological” refers to little more than the obvious fact that behavior begins, one way or another, in the brain, and differences in behavior begin, one way or another, in the genome.
Kendler’s further analysis depends on the concept of biological coherence: “by coherence, I mean the genes whose altered expression or structure is indexed by the detected common or rare genetic variants or [copy number variations] tell a sensible biological story.” He describes four “scenarios” with different degrees of coherence (It would seem that four per se, or really breaking coherence down into categories at all, is more a matter of explanatory convenience than anything else. The four scenarios are basically no coherence, two levels of middling coherence, and high coherence.)
When all the analysis is complete, I expect that Kendler’s appeal to coherence and my phenotypic null hypothesis will wind up pointing in more or less the same direction. In the meantime, however, I will express one concern: How exactly is coherence to be detected? Kendler’s examples come exclusively from widely recognized medical disorders: Crohn’s disease, Parkinson’s diseases, multiple sclerosis, and Alzheimer’s disease. These examples are ultimately too easy: no one has ever mistaken Alzheimer’s disease for a lifestyle. The judges of coherence seem to be the working geneticists who study the syndromes, who—unsurprisingly-- are unanimous in concluding that the genetics of their respective disorders are headed in a coherent direction. Scientists always think this way, and I for one will be more convinced that the coherence criterion is going to work when someone identifies a meaningful domain of behavior where coherence doesn’t seem to obtain. Kendler does not offer a suggestion.
In the consideration of the (no coherence) Scenario 1, Kendler concludes, “I consider Scenario 1 to be improbable. Although psychiatric disorders are likely etiological heterogeneous, and some of the coherence may be emergent at levels in the mind-brain system too high for us to now study, the chances that no meaningfully connectivity between variants will emerge seems low.” For the record, it doesn’t seem so low to me, given the history of biological psychiatry up to this point, and an exhaustive review of the personality literature has convinced me that Scenario 1 is already here for normal personality. Once again, any attempt at a theory of biological explanation in psychiatry has to set a marker on the left edge of the domain, identifying something that is not biological, if it wants to be taken at its word about things that are.
Conclusion: The Meaning of Nature
When one starts to enumerate the great theoretical questions in the history of psychiatry, it is striking how many of them depend crucially on the nature question and questions of mechanism, and how few of them hang on Galtonian considerations of genetic epidemiology:
● Diagnostic realism and natural kinds vs. social construction
● The organic-functional distinction
● Mental illness as “myth” and medical reality
● Disease models of addiction
● Treatment with pharmaceuticals as opposed to talk therapy
● Considerations of moral responsibility in the legal system
● The possibility of objective biological diagnosis of psychopathology
Galton’s pleasing jingle of nature and nurture has become so familiar that it is easy to forget that nature in this sense, as what is inherited as opposed to inculcated, is not even its oldest or most common meaning in connection to human behavior. Neither is it the most profound. The other meaning of nature, as in the phrase human nature owes less to Galton, and more to another late nineteenth century interpreter of complex human biology, Freud. To ask about human nature, at the species level, is to wonder whether humans in general are universally endowed or burdened with desires or capacities that are beyond their willful control. At the individual level, to speak of a person’s nature is to consider the possibility, once again, that some personal desires are not to be explained in terms of conscious intentions and observable life narratives, but instead by inner mechanisms of which a person may be only dimly aware. The relation between these levels-- the expression of primitive species-typical desires in the presumably civilized minds of modern individuals-- is the central concern of Freudianism.
Nature vs. nurture in the Galtonian, genetic epidemiological sense has little to do with the determination of human nature at either the species or the individual level, and questions of human nature remain undecided once the Galtonian questions have been answered. Another important usage of the word nature is important in this context. One of the attractions of natural explanations of human behavior is that they offer a route to natural science, a way out of the perennial methodological swamp of the behavioral sciences. Freud was not immune to this temptation. In the famous opening to his never-completed “Project for a Scientific Psychology,” he wrote:
The intention is to furnish a psychology that shall be a natural science: that is, to represent psychical processes as quantitatively determinate states of specifiable material particles, thus making these psychical processes perspicuous and free of contradiction.’ (Freud, 1895)
In the contrast between Freud and the unexamined materialism of modern biological psychiatry we can see both horns of the dilemma that has plagued psychiatry from the start: Indulge a high-level, humanistic model of causation and explanation, paying a price in methodological rigor, or indulge an exclusively reductionist view grounded in the natural sciences, paying a price in psychological relevance. Galtonian questions of nature and nurture are not irrelevant to this dilemma so much as orthogonal to it. Natural science explanations of human behavior must recognize that genetic variability does not guarantee the success of bottom-up explanation, or risk losing their relevance to human suffering as it is actually experienced. Psychological theories must remain compatible with genetic variation, or risk losing their relevance to the world of empirical science.
References
Beam, C. R., & Turkheimer, E. (2013). Phenotype–environment correlations in longitudinal twin models. Development and psychopathology, 25(1), 7-16.
Charney, E. (2012). Behavior genetics and postgenomics. Behavioral and Brain Sciences, 35(05), 331-358.
Chua, S. E., & McKenna, P. J. (1995). Schizophrenia--a brain disease? A critical review of structural and functional cerebral abnormality in the disorder.The British Journal of Psychiatry, 166(5), 563-582.
Crow, T. J. (2008). The emperors of the schizophrenia polygene have no clothes. Psychological medicine, 38(12), 1681.
D’Onofrio, B. M., Turkheimer, E., Emery, R. E., Harden, K. P., Slutske, W. S., Heath, A. C., ... & Martin, N. G. (2007). A genetically informed study of the intergenerational transmission of marital instability. Journal of Marriage and Family, 69(3), 793-809.
Faraone, S. V., & Tsuang, M. T. (1985). Quantitative models of the genetic transmission of schizophrenia. Psychological Bulletin, 98(1), 41.
Freud, S. (1895). Project for a scientific psychology. In J. Strachey, ed. and trans., The standard edition of the complete psychological works of Sigmund Freud, 24 vols. London: Hogarth Press, 1953-1974. 1:281–391.
Galton, F. (1875). English men of science: Their nature and nurture. D. Appleton.
Galton, F. (1876). The history of twins, as a criterion of the relative powers of nature and nurture. The Journal of the Anthropological Institute of Great Britain and Ireland, 5, 391-406.
Gottesman, I. I. & Shields, J. (1962). Schizophrenia and Genetics: A Twin Study Vantage Point. New York: Academic Press.
Guze, S. B. (1989). Biological psychiatry: is there any other kind? Psychological medicine, 19(2), 315-323.
Hatemi, P. K., Funk, C. L., Medland, S. E., Maes, H. M., Silberg, J. L., Martin, N. G., & Eaves, L. J. (2009). Genetic and environmental transmission of political attitudes over a life time. Journal of Politics, 71(3), 1141-56.
Heinrichs, R. W. (1993). Schizophrenia and the brain: Conditions for a neuropsychology of madness. American Psychologist, 48(3), 221.
Henn, F. A., & Nasrallah, H. A. (1982). Schizophrenia as a brain disease. Oxford University Press.
Johnson, D. L. (1989). Schizophrenia as a brain disease: Implications for psychologists and families. American Psychologist, 44(3), 553.
Joseph, J. (2004). The gene illusion: Genetic research in psychiatry and psychology under the microscope. Algora Publishing.
Kendler, K. S. (2013). What psychiatric genetics has taught us about the nature of psychiatric illness and what is left to learn. Molecular psychiatry.
Kety, S. (1974). From rationalization to reason. American Journal of Psychiatry, 131, 957-963.
Lewontin, R. C., Rose, S. & Kamin, L. J. (1984). Not in our genes: Biology, ideology and human nature. New York: Pantheon Books.
Lidz, T., & Blatt, S. (1983). Critique of the Danish-American studies of the biological and adoptive relatives of adoptees who became schizophrenic.American Journal of Psychiatry, 140(4), 426-35.
Lipowski, Z. J. (1989). Psychiatry: mindless or brainless, both or neither?.Canadian journal of psychiatry. Revue canadienne de psychiatrie, 34(3), 249.
Loehlin, J. C., & Martin, N. G. (2013). General and supplementary factors of personality in genetic and environmental correlation matrices. Personality and Individual Differences, 54(6), 761-766.
Meehl, P. E. A critical afterword. In I. I. Gottesman & J. Shields, Schizophrenia and genetics. New York: Academic Press, 1972.
Plomin, R., Corley, R., DeFries, J. C., & Fulker, D. W. (1990). Individual differences in television viewing in early childhood: Nature as well as nurture.Psychological Science, 1(6), 371-377.
Rende, R. D., Plomin, R., & Vandenberg, S. G. (1990). Who discovered the twin method?. Behavior genetics, 20(2), 277-285.
Schwarz, S. C. (1991) Genetics of Schizophrenia: A status report. In A. Tasman & S. M. Goldfinger (Eds.), American Psychiatric Press Review of Psychiatry, Volume 10. Washington, DC: American Psychiatric Press, 1991, 79-97.
Szasz, T. S. (1960). The myth of mental illness. American Psychologist, 15(2), 113.
Turkheimer, E. (1998). Heritability and biological explanation. Psychological Review, 105(4), 782.
Turkheimer, E. (2000). Three laws of behavior genetics and what they mean.Current Directions in Psychological Science, 9(5), 160-164.
Turkheimer E, Harden KP. (In press). Behavior genetic research methods: Testing quasi -causal hypotheses using multivariate twin data. In Handbook of Research Methods in Personality and Social Psychology, ed. HT Reis, CM Judd. Cambridge Univ. Press. 2nd ed.
Turkheimer, E., Horn, E. H. & Pettersson, E. (in press). A phenotypic null hypothesis for the genetics of personality. Annual Review of Psychology.
Visscher, P. M., Hill, W. G., & Wray, N. R. (2008). Heritability in the genomics era—concepts and misconceptions. Nature Reviews Genetics, 9(4), 255-266.
Waller, N. G., Kojetin, B. A., Bouchard, T. J., Lykken, D. T., & Tellegen, A. (1990). Genetic and environmental influences on religious interests, attitudes, and values: A study of twins reared apart and together. Psychological Science,1(2), 138-142.
[1] As Billy Martin put it, “Well, you got your mules and you got your racehorses, and you can kick a mule in the ass all you want, and he's still not gonna be a racehorse.”